کشش ماهیچه
کشش ماهیچه (به انگلیسی: Muscle Contraction) (یا انقباض ماهیچه)، فعال سازی مناطق مولد کشش در تارهای ماهیچهایست.[۱][۲] در فیزیولوژی، انقباض ماهیچه لزوماً به معنای کوتاه شدن ماهیچه نیست، چرا که کشش ماهیچه ممکن است بدون تغییر طول ماهیچه رخ دهد، مثل زمانی که کتاب سنگین یا دمبل، در موقعیت یکسانی نگه داشته شود.[۱] پایان انقباض ماهیچهای، شلشدن ماهیچهای (Muscle Relaxation) است، که بازگشت تارهای ماهیچهای به حالتیست که در آن کشش کمی تولید میگردد.[۱]


انقباضات ماهیچهای را میتوان براساس دو متغیر توصیف کرد: طول و کشش.[۱] کشش ماهیچهای را ایزومتریک گویند اگر کشش ماهیچه تغییر کند اما طول آن بدون تغییر باقی بماند.[۱][۳][۴][۵] در مقایسه، کشش ماهیچهای را ایزوتونیک گویند اگر کشش ماهیچه در طول کشش بدون تغییر باقی بماند.[۱][۳][۴][۵] اگر طول ماهیچه کمتر شود، کشش را هممرکز (concentric) گفته؛[۱][۶] و در صورتی که طول ماهیچه طی کشش بلندتر شود، کشش را غیر-هممرکز (eccentric) مینامند. در حرکات طبیعی که باعث جنبش جانوران میگردد، کششهای ماهیچهای اغلب چندوجهی بوده، یعنی ماهیچهها قادرند برحسب زمان، طول و کشش تولیدیشان را تغییر دهند.[۷] بنابراین، احتمال این که نه طول و نه کشش ماهیچه، طی فعالیتهای حرکتی جانور تغییر نکند، اندک است.
در مهرهداران، کشش ماهیچههای اسکلتی نوروژنیک است، یعنی نیازمند ورودی سیناپسی از نورونهای حرکتی جهت تولید کششهای ماهیچهای میباشند. یک نورون حرکتی واحد، قادر به عصبدهی به چندین تار ماهیچهایست، بنابر این باعث کشش همزمان تارهایی که به آنها متصل شده، میگردد. پس از عصبدهی، فیلامانهای پروتئینی درون تار ماهیچه اسکلتی، بر روی یکدیگر لغزیده تا ایجاد کشش یا انقباض کنند؛ این فرایند توسط نظریه لغزش فیلامانی توضیح داده میشود. کشش تولید شده را میتوان براساس فرکانس پتانسیل عمل به صورت تکانش یا کشش ناگهانی (twitch)، جمعبندی (حالتی که تکانشی قبل از شل شدن کامل تکانش قبلی ایجاد میگردد)، یا تشنج (تتانوس، وضعیتی که در کزاز پیش میآید) توصیف نمود. در عضلات اسکلتی، کشش عضلانی هنگامی به بیشترین مقدر خود میرسد که عضله منقبض شده تا بر اساس رابطه کشش-طول، به طول میانه خود برسد.
برخلاف ماهیچههای اسکلتی، کشش ماهیچههای صاف و قلبی به صورت میوژنیک میباشند (به این معنا که این انقباضات توسط خود سلولهای عضلانی قلبی یا صاف آغاز میگردد، نه این که از رویداد بیرونی چون تحریک عصبی آغاز شوند)، گرچه که تحریکات دستگاه عصبی خودمختار نیز قادر به تعدیلشان میباشد. سازوکارهای کششی در این بافتهای ماهیچهای مشابه با بافتهای ماهیچههای اسکلتیست.
انواع
ویرایشانقباض ماهیچه اسکلتی
ویرایشبا رسیدن پیام عصبی به سلول عضلانی یا همان سارکوپلاسم (به انگلیسی: sarcoplasm) و غیر متعادل کردن غشای آن به نام سارکولما (به انگلیسی: sarcolemma) از نظر بار الکتریکی، این عدم تعادل از طریق فرو رفتگیهایی در سارکولما به نام لوله تی (به انگلیسی:t tubule) به عمق سارکولما و در محلی که رتیکولومهای سارکوپلاسمی حضور دارند نفوذ میکند و سبب آزاد شدن یونهای کلسیم از این رتیکولومهای سارکوپلاسمی میشود. با افزایش آزاد شدن یونهای کلسیم از رتیکولومهای سارکوپلاسمی (به انگلیسی: Sarcoplasmic reticulum) درون سارکولما به درون تارچه یا میوفیبریلهای عضلانی تعداد بیشتری از این یونها به درون فضای سارکومر در درون میوفیبریل میروند که در آنجا به پروتئینهای تروپونین (به انگلیسی: troponin) که بر روی رشتههای تروپومیوزین (به انگلیسی: tropomyosin) که خود بر روی رشتههای پروتئینی اکتین (به انگلیسی: actin) قرار دارند چسبیده و پس از واکنش با آنها منجر به حرکت کردن آنها از سر جایشان میشوند که این حرکت منجر به آزاد شدن محل اتصال رشتههای پروتئینی میوزین (به انگلیسی: myosin) به اکتین میشود. پس از آزاد شدن محل اتصال زائدههای موی شکلی (به انگلیسی: bulbous head) که بر روی رشتههای پروتئینی میوزین وجود دارند به درون محلهای اتصال روی رشتههای پروتئینی اکتین فرورفته و اتصالی به نام پل متقاطع (به انگلیسی:cross bridge) را تشکیل میدهند. این حرکت منجر به آزاد شدن فسفاتهایی که بر روی زائدههای میوزین قرار داشتهاند میشوند، با آزاد شدن فسفاتها آدنوزین دی فسفات (ADP)هایی که تاکنون در سر این زائدهها ذخیره شده بودند واکنش داده و انرژی مورد نیاز برای حرکت این زائدهها را در یک جهت و به میزان ۴۵ درجه را فراهم میآورند با حرکت این زائدهها رشتههای اکتینی که به این زائدهها ی میوزینی متصل هستند نیز حرکت میکنند. با حرکت رشتههای اکتینی و با کوتاه شدن نوار اچ (به انگلیسی: H band) عضله منقبض میشود این زنجیره از فرایندها تا حدی تکرار میشود که عضله به میزان انقباض لازم برسد. پس از هر حرکت یک آدنوزین تریفسفات (ATP) جدید به سوی سر زائدههای میوزینی آمده و در همین هنگام این زائدهها از رشتهٔ اکتین جدا میشوند. آدنوزین تریفسفات در سر زائدههای میوزینی توسط آنزیم ای تی پاز (به انگلیسی: atpase) به آدنوزین دی فسفات و یک فسفر (Pi) تجزیه میشود، اکنون زائدههای میوزینی آماده برای انقباض بعدی عضلانی میباشند. پس از پایان یافتن هر انقباض یونهای کلسیم به رتیکولومهای سارکوپلاسمی (به انگلیسی: Sarcoplasmic reticulum) ای در خارج از میوفیبریل که از آنها به هنگام رسیدن پیام عصبی آزاد شده بودند بازمیگردند.
-
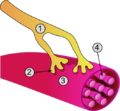 در نگارهٔ بالا: ۱. آکسون ۲. پایانه آکسون ۳. سارکومر ۴. تارچه میباشند.
در نگارهٔ بالا: ۱. آکسون ۲. پایانه آکسون ۳. سارکومر ۴. تارچه میباشند. -
 فضای درون سارکومر
فضای درون سارکومر


جستارهای وابسته
ویرایشمنابع
ویرایش- ↑ ۱٫۰ ۱٫۱ ۱٫۲ ۱٫۳ ۱٫۴ ۱٫۵ ۱٫۶ Widmaier, Eric P.; Raff, Hersel; Strang, Kevin T. (2010). "Muscle". Vander's Human Physiology: The Mechanisms of Body Function (12th ed.). New York, NY: McGraw-Hill. pp. 250–291. ISBN 978-0-321-98122-6.
- ↑ Silverthorn, Dee Unglaub (2016). "Muscles". Human Physiology: An Integrated Approach (7th ed.). San Francisco, CA: Pearson. pp. 377–416. ISBN 978-0-321-98122-6.
- ↑ ۳٫۰ ۳٫۱ Aidley, David J. (1998). "Mechanics and energetics of muscular contraction". The Physiology of Excitable Cells (4th ed.). New York, NY: Cambridge University Press. pp. 323–335. ISBN 978-0-521-57421-1.
- ↑ ۴٫۰ ۴٫۱ Sircar, Sabyasachi (2008). "Muscle elasticity". Principles of Medical Physiology (1st ed.). New York, NY: Thieme. pp. 113. ISBN 978-1-58890-572-7.
- ↑ ۵٫۰ ۵٫۱ Bullock, John; Boyle, Joseph; Wang, Michael B. (2001). "Muscle contraction". NMS Physiology. Vol. 578 (4th ed.). Baltimore, Maryland: Lippincott Williams and Wilkins. pp. 37–56.
- ↑ Kumar, Shrawan (2008). "Introduction and terminology". In Shrawan Kumar (ed.). Muscle strength (1st ed.). Boca Raton, FL: CRC Press. p. 113. ISBN 978-0-415-36953-4.
- ↑ Biewener, Andrew A. (2003). "Muscles and skeletons: The building blocks of animal movement". Animal Locomotion. Oxford Animal Biology Series. New York, NY: Oxford University Press. pp. 15–45. ISBN 978-0-19-850022-3.
- مشارکتکنندگان ویکیپدیا. «Muscle Contraction». در دانشنامهٔ ویکیپدیای انگلیسی.