ماهیچه
ماهیچه یا عضله (به انگلیسی: Muscule)، نوعی بافت بدن است که برای حرکت بدن استفاده میشود. ماهیچهها انرژی شیمیایی مواد غذایی را به انرژی مکانیکی تبدیل میکنند، حرکت بدن از انقباض و انبساط ماهیچهها حاصل میشود. ماهیچهها به نسبت شکل و اندازهای که دارند، تقسیمبندی میشوند، در بدن سه نوع ماهیچه وجود دارد:
- ماهیچه اسکلتی (مخطط) که در افراد معمولی حدود ۴۰٪ وزن بدن را تشکیل میدهد.
- ماهیچه صاف مانند ماهیچههای خودکار دیواره رگها، روده و معده که حدود ۱۰٪ از بدن را تشکیل میدهد.
- ماهیچه قلبی مخطط است ولی غیرارادی است. ما فقط یک عضله مخطط منشعب و غیرارادی در بدن داریم که عضله قلب است.
| Muscle | |
|---|---|
 | |
| جزئیات | |
| ساخته از | میانپوست |
| دستگاه | دستگاه عضله اسکلتی انسان |
| شناسهها | |
| لاتین | musculus |
| MeSH | D009132 |
| TA2 | 1975, 1994 |
| FMA | 5022 30316، 5022 |
ساختمان بافتی تارچههای ماهیچه
ویرایشدر ساختمان بافتی تارچههای ماهیچهای در هر سارکومر دو نوع میوفیلامان (میوفیلامنت) قطور و نازک وجود دارد. ترکیب شدن و جابهجایی پروتئینهایی که در این میوفیلامانها وجود دارند، اساس مولکولی انقباض ماهیچهای و کوتاه شدن سارکومرها را تشکیل میدهند. هر یک از میوفیلامانهای قطور که در بخش میانی سارکومر قرار دارند از صدها مولکول پروتئینی به نام میوزین ساخته شدهاند. هر مولکول میوزین شبیه میلهٔ نازکی با یک سر کروی است که با زاویهٔ خاصی به صورت یک پل عرضی به یک میوفیلامان نازک میچسبد. میوفیلامانهای نازک دارای سه نوع پروتئیناند: پروتئین اصلی آنها اکتین نام دارد که مولکولهای آن کروی و کوچک است که به صورت یک زنجیرهٔ دوتایی قرار گرفتهاند. دو پروتئین دیگر تروپومیوزین و تروپونین نام دارند.
تروپونین دارای سه زیرواحد است. این زیرواحدها عبارتند از تروپونین I، تروپونین T که به تروپومیوزین متصل میشود و تروپونین C که به کلسیم متصل میشود.
تروپومیوزین به صورت مولکولهای دراز و رشتهای است و هر یک از این مولکولها روی چند مولکول اکتین را میپوشاند.
مولکولهای تروپونین C که در طول رشتههای تروپومیوزین چسبیدهاند، با یونهای کلسیم میل ترکیبی شدید دارند و اتصال آنها با یونهای کلسیم باعث شروع مکانیسم مولکولی انقباض میشود.
در نهایت ماهیچه دارای توالیهای سارکومری است که پروتئینهای موجود در این توالیها میتوانند تارهای ماهیچهای را به انقباض وادار نمایند.
فیزیولوژی
ویرایش
در حال استراحت ماهیچه، مجموعهٔ مولکولهای تروپومیوزین و تروپونین از ترکیب مولکولهای اکتین و میوزین جلوگیری میکنند.
بر اثر تحریک عصبی و اثر استیلکولین بر گیرندههای سلول ماهیچهای، کلسیم از شبکه سارکوپلاسمی آزاد میشود. وقتی کلسیم بر تروپونین C اثر کند، تغییری در آرایش مولکولها پدید میآید که باعث جدا شدن تروپومیوزین از اکتین میشود. حال اکتین آزاد شده و میوزین میتواند به آن متصل شود و باعث پلهای عرضی متناوب به مولکولهای اکتین و لغزاندن میوفیلامانهای نازک بر روی میوفیلامانهای قطور شود. به این ترتیب که سرهای فعال مولکولهای میوزین با زاویهٔ معینی به مولکولهای اکتین میچسبد و آنها را مانند یک چرخ دندانهدار جابهجا میکنند. این تغییرات باعث کوتاه شدن سارکومرها و نزدیک شدن خطوط Z به یکدیگر و در نتیجه کوتاه شدن طول ماهیچه میشود.
در هنگام انقباض ماهیچه پلهای عرضی میوفیلامانهای قطور پس از اتصال به میوفیلامانهای نازک و حرکت دادن آنها از میوفیلامانهای نازک جدا شده و دوباره در نقطهٔ دورتری به آن متصل میشوند. انرژی لازم برای این جابهجایی از مصرف آدنوزین تریفسفات (ATP) به وسیلهٔ سر فعال میوزین حاصل میشود.
انقباض ماهیچه اسکلتی
ویرایشبا رسیدن پیام عصبی به سلول ماهیچهای یا همان سارکوپلاسم (به انگلیسی: Sarcoplasm) و غیر متعادل کردن غشای آن به نام سارکولما (به انگلیسی: Sarcolemma) از نظر بار الکتریکی، این عدم تعادل از طریق فرو رفتگیهایی در سارکولما به نام لوله تی (به انگلیسی:t tubule) به عمق سارکولما و در محلی که شبکههای سارکوپلاسمی حضور دارند نفوذ میکند و سبب آزاد شدن یونهای کلسیم از این شبکهها میشود. با افزایش آزاد شدن یونهای کلسیم از شبکههای سارکوپلاسمی درون سارکولما به درون تارچه یا میوفیبریلهای عضلانی تعداد بیشتری از این یونها به درون فضای سارکومر در درون میوفیبریل میروند که در آنجا به پروتئینهای تروپونین که بر روی رشتههای تروپومیوزین (به انگلیسی: tropomyosin) که خود بر روی رشتههای پروتئینی اکتین قرار دارند چسبیده و پس از واکنش با آنها منجر به حرکت کردن آنها از سر جایشان میشوند که این حرکت منجر به آزاد شدن محل اتصال رشتههای پروتئینی میوزین به اکتین میشود. پس از آزاد شدن محل اتصال زائدههای موی شکلی (به انگلیسی: bulbous head) که بر روی رشتههای پروتئینی میوزین وجود دارند به درون محلهای اتصال روی رشتههای پروتئینی اکتین فرورفته و اتصالی به نام پل متقاطع (به انگلیسی:cross bridge) را تشکیل میدهند.
این حرکت سبب آزاد شدن فسفاتهایی که بر روی زائدههای میوزین قرار داشتهاند میشوند، با آزاد شدن فسفاتها آدنوزین دیفسفات (ADP)هایی که تاکنون در سر این زائدهها ذخیره شده بودند واکنش داده و انرژی مورد نیاز برای حرکت این زائدهها در یک جهت و به میزان ۴۵ درجه را فراهم میآورند. با حرکت این زائدهها رشتههای اکتینی که به این زائدههای میوزینی متصل هستند نیز حرکت میکنند. با حرکت رشتههای اکتینی و با کوتاه شدن نوار اچ (به انگلیسی: H band) عضله منقبض میشود. این زنجیره از فرایندها تا حدی تکرار میشود که ماهیچه به میزان انقباض لازم برسد. پس از هر حرکت یک آدنوزین تریفسفات جدید به سوی سر زائدههای میوزینی آمده و در همین هنگام این زائدهها از رشتهٔ اکتین جدا میشوند. آدنوزین تریفسفات در سر زائدههای میوزینی توسط آنزیم ایتیپاز به آدنوزین دیفسفات و یک فسفر تجزیه میشود، اکنون زائدههای میوزینی آماده برای انقباض بعدی عضلانی هستند.
پس از پایان یافتن هر انقباض، یونهای کلسیم به شبکههای سارکوپلاسمی در خارج از میوفیبریل که از آنها به هنگام رسیدن پیام عصبی آزاد شده بودند بازمیگردند.
-
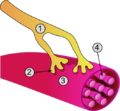 در نگارهٔ بالا: ۱. آکسون ۲. پایانه آکسون ۳. سارکومر ۴. تارچه هستند.
در نگارهٔ بالا: ۱. آکسون ۲. پایانه آکسون ۳. سارکومر ۴. تارچه هستند. -
 فضای درون سارکومر
فضای درون سارکومر


جستارهای وابسته
ویرایشمنابع
ویرایشبروس آلبرتس، (۱۳۸۵)، مبانی زیستشناسی سلولی، ترجمهٔ بهرام میرحبیبی، حسین بهاروند، پروانه فرزانه، داوود صبور، علی شیخیان، مرضیه ابراهیمی، محمد پاکزاد، نرگس زارع، شبنم زرعی مرادی، سیدمحمود هاشمی، مؤسسه فرهنگی انتشاراتی تابش اندیشه، ص. ص٫، شابک ۹۶۴-۹۵۶۴۷-۲-۱